The Phoenix Islands Protected Area (PIPA) is the largest marine protected area in the Pacific Ocean. As part of the permitting process which enables us to work on Nikumaroro, we are required to submit a report to PIPA at the end of each expedition, describing what we did, what we saw, and including video and still images of the island and its environment. The report was delivered in person by Ric Gillespie on June 23, 2011, in Tarawa, and is published below. There are also numerous appendices which are published in the Analysis section.
Appendix H
An Analysis of Marine Mollusk Shells
From the Seven Site, Nikumaroro Atoll,
Phoenix Islands, Republic of Kiribati
June, 2011
Judith R. Amesbury
Micronesian Archaeological Research Services
Guam
With an appendix by
Dr. Katherine Szabó, QEII Research Fellow
School of Earth and Environmental Sciences
University of Wollongong
Australia
Prepared for The International Group for Historic Aircraft Recovery
Introduction
In December 2010, Micronesian Archaeological Research Services (MARS) was contracted by The International Group for Historic Aircraft Recovery (TIGHAR) to analyze marine mollusk shells collected from the Seven Site on Nikumaroro Atoll in the Phoenix Islands, Republic of Kiribati. MARS agreed to analyze the shells in the same ways in which we analyze other archaeological assemblages, but with special attention to details, such as breakage patterns of the shells, that might help to determine if these mollusks had been collected and eaten by a Pacific Islander or by a non-Pacific Islander.
Methods
MARS received five bags of shells from the Seven Site: two from Feature C (one collected in 2001 and one collected in 2010), two from Feature D (both collected in 2010), and one from Feature SL (also collected in 2010). These were not samples of the features; they were complete collections of the mollusk features.
In addition MARS received two bags that were grab samples. One was from the lagoon beach and the other was from a berm that fronts the lagoon shore. Both of these samples were collected near the Seven Site for comparison with the shells in the cultural features (C, D, and SL).
Faunal analysts quantify animal remains in at least three ways: 1) by the number of identified specimens (NISP); 2) by the weight of the remains, usually in grams (g); and 3) by the minimum number of individuals (MNI). The MNI of a mollusk species is determined by counting only whole shells and fragments with a distinctive feature to determine how many individual animals were present. For example, for bivalves the analyst counts the number of whole shells and fragments with hinges present and divides by two, since bivalves have two shells. For gastropods, the analyst counts only the spires or only the apertures or some other distinctive feature, such as the stromboid notch on strombids. Grayson (1984) explains the three methods and discusses the advantages of each. Amesbury decided to use all three methods in hopes of gaining the maximum information from the shells.
In the MARS laboratory, the shells were sorted to the lowest possible taxon (family, genus, or species) and counted and weighed. Because some of the shells are broken and almost all are faded of their colors, sometimes the species cannot be determined. In scientific nomenclature, if a species is unknown, the abbreviation “sp.” is used after the genus name. If there is more than one unknown species within a single genus, the abbreviation “spp.” is used after the genus name. If the analyst is unsure of the species, but wants to indicate what species it is similar to, the abbreviation for the word compare, “cf.”, is used between the genus and species names.
Whole shells and fragments were counted and weighed separately. MNI was determined for the cultural features as explained above. Because the vast majority of the shells in the cultural features are from a certain genus of Venus clam, Gafrarium, the shells of that genus were further examined as to the ways in which they were broken.
MNI was not determined for the grab samples, because they are not quantitative samples. The grab sample from the berm fronting the lagoon shore was not completely sorted. Because it was composed of small and eroded shells and pieces of coral, the sorting was very time consuming, and the last 50 grams of shell and coral fragments were left unsorted.
All of the bags were analyzed separately in order that no information would be lost. After the separate analysis, the results of the two bags from Feature C were combined as were the results of the two bags from Feature D. The shells were not combined.
Results
Identification and Quantification
The results of the initial analysis of each bag are found in Appendix A. The results of the analysis by feature with MNI are shown here in Tables 1-3.
Table 1. Minimum number of individuals (MNI) and number and weight in grams of whole shells and fragments from Feature C.
FAMILY
GENUS
SPECIES
MNI
Wh. #
Wt.(g)
Fr. #
Wt.(g)
Total Wt.
Percent of Wt.
Tellinidae
>
Quidnipagus
palatam
1
1
1.0
>
1.0
0.11
Tridacnidae
Tridacna
maxima
1
1
45.2
45.2
4.76
Veneridae
Gafrarium
tumidum
123
173
643.4
146
241.3
884.7
93.26
Bivalves
125
174
688.6
147
242.3
930.9
98.13
Cypraeidae
Cypraea
sp.
1
1
1.2
1.2
0.13
Conidae
Conus
sp.
1
1
2.6
2.6
0.27
Turbinidae
Turbo
sp.
1
4
13.9
13.9
1.47
Gastropods
3
6
17.7
17.7
1.87
TOTALS
128
174
688.6
153
260.0
948.6
100.00
Table 2. Minimum number of individuals (MNI) and number and weight in grams of whole shells and fragments from Feature D.
FAMILY
GENUS
SPECIES
MNI
Wh. #
Wt. (g)
Fr. #
Wt. (g)
Total Wt.
Percent of Wt.
Cardiidae
Fragum
fragum
45
45
13.7
13.7
0.73
Mytilidae
1
1
0.1
0.1
0.01
Veneridae
Gafrarium
tumidum
269
114
416.7
1129
1207.1
1623.8
86.46
Bivalves
315
159
430.4
1130
1207.2
1637.6
87.19
Cerithiidae
9.7
0.52
Cerithium
nodulosum
1
1
5.8
Other
spp. (at least 2)
6
2
0.5
5
3.4
Conidae
Conus
spp. (at least 3)
9
8
45.9
4
4.5
50.4
2.68
Cypraeidae
Cypraea
spp. (at least 3)
29
20
102.9
18
59.9
162.8
8.67
Muricidae
2
2
3.4
3.4
0.18
Neritidae
Nerita
sp.
1
1
0.5
0.5
0.03
Strombidae
2
2
1.6
1.6
0.09
Turbinidae
Turbo
sp.
2
5
12.1
12.1
0.64
Gastropods
52
30
149.3
38
91.2
240.5
12.81
TOTALS
367
189
579.7
1168
1298.4
1878.1
100.00
Table 3. Minimum number of individuals (MNI) and number and weight in grams of whole shells and fragments from Feature SL.
FAMILY
GENUS
SPECIES
MNI
Wh. #
Wt. (g)
Fr. #
Wt. (g)
Total Wt.
Percent
of Wt.
Cardiidae
Fragum
fragum
2
2
0.8
0.8
0.24
Tridacnidae
Tridacna
maxima
1
1
5.6
5.6
1.68
Veneridae
321.0
96.02
Gafrarium
tumidum
32
31
162.9
46
145.5
Pitar
prora
2
2
12.6
Bivalves
37
36
181.9
46
145.5
327.4
97.94
Turbinidae
Turbo
sp.
1
2
6.8
6.8
2.03
Unidentified gastropods
0
1
0.1
0.1
0.03
Gastropods
1
3
6.9
6.9
2.06
TOTALS
38
36
181.9
47
152.4
334.3
100.00
Total weight of all shells from the three cultural features is 3,161 grams or about 7 pounds. Feature D is the largest feature with almost two kilograms of shell. Feature C is only about half that size with almost one kilogram. Feature SL is the smallest with about one-third of a kilogram.
The three cultural features differ as to the number of species present. As might be expected, the largest feature, Feature D, has the most species (at least sixteen species belonging to ten families). Feature C has six species belonging to six families, and Feature SL has five species belonging to four families, plus a small fragment of an unidentified gastropod, which may or may not be a different species or family.
All of the cultural features have a greater weight of bivalves than gastropods, and in all the features the most abundant genus by weight, NISP and MNI is Gafrarium. It appears that the person (or people) who collected the mollusks was targeting that genus. Neither of the comparative grab samples (from the lagoon beach and the berm) have such high counts and weights of Gafrarium
The percentage of the total weight of Gafrarium in the lagoon beach grab sample is slightly less than 25 percent (Table 4), and there is almost no Gafrarium present in the berm grab sample (Table 5). But the percentages by weight of Gafrarium in the three cultural features are 93 percent (Feature C), 86 percent (Feature D), and 92 percent (Feature SL).
Table 4. Number and weight in grams of whole shells and fragments from the lagoon beach grab sample.
FAMILY
GENUS
SPECIES
Wh. #
Wt. (g)
Fr. #
Wt. (g)
Total Wt.
Percent
of Wt.
Arcidae
Barbatia
cf. amygdalumtostum
1
5.2
5.2
1.06
Cardiidae
Fragum
fragum
1
0.2
1
2.2
2.4
0.49
Chamidae
Chama
cf. iostoma
3
36.0
36.0
7.37
Lucinidae
Ctena
bella
2
4.1
4.1
0.84
Tellinidae
Quidnipagus
palatam
3.8
3.8
0.78
Tridacnidae
222.6
45.57
Tridacna
maxima
4
46.4
2
51.9
Tridacna
spp. (at least 2)
3
124.3
Veneridae
214.4
43.89
Gafrarium
tumidum
28
120.6
Pitar
prora
10
78.1
1
2.0
Lioconcha
cf. castrensis
2
13.7
Bivalves
52
308.1
7
180.4
488.5
100.00
Gastropods
0
0
0
0
0
0.00
TOTALS
52
308.1
7
180.4
488.5
100.00
Table 5. Number and weight in grams of whole shells and fragments from the grab sample from the berm that fronts the lagoon shore.
FAMILY
GENUS
SPECIES
Wh. #
Wt. (g)
Fr. #
Wt. (g)
Total Wt.
Percent of Wt.
Cardiidae
Fragum
fragum
333
63.0
19
2.8
65.8
82.46
Tellinidae
probably 2 species
2
0.2
0.2
0.25
Veneridae
Gafrarium
tumidum
2
0.5
0.5
0.63
Unidentified bivalves
10
1.0
1.0
1.25
Bivalves
333
63.0
33
4.5
67.5
84.59
Cerithiidae
at least 2 species
41
7.8
27
3.8
11.6
14.53
Strombidae
Strombus
sp.
1
0.7
0.7
0.88
Gastropods
41
7.8
28
4.5
12.3
15.41
TOTALS*
374
70.8
61
9.0
79.8
100.00
* Included with the shells on this table are 169 small pieces of coral weighing 23.6 grams. In addition to the above shells and the accompanying 23.6 grams of coral, there are 50 grams of unsorted shells and coral pieces with approximately the same make-up as the shells and corals that were counted and weighed. The total weight of shells and coral fragments from the berm grab sample is therefore 153.4 grams.
The Gafrarium shells have been identified on the tables as Gafrarium tumidum. It is possible that a small number of the shells are a very similar species, Gafrarium pectinatum. It is difficult to distinguish between the two species, especially if the shells are not whole and mature specimens. Shells of mature G. tumidum are larger and thicker than shells of G. pectinatum. Shells of G. tumidum are blunter anterior to posterior, while G. pectinatum shells are more elongated anterior to posterior. Although the sculpture is similar on both species, it is somewhat more pronounced on G. tumidum.
Breakage Patterns
The three cultural features (C, D, and SL) all exhibit the same patterns of breakage of the Gafrarium (Tables 6-8), but the frequencies of the breakage patterns differ.
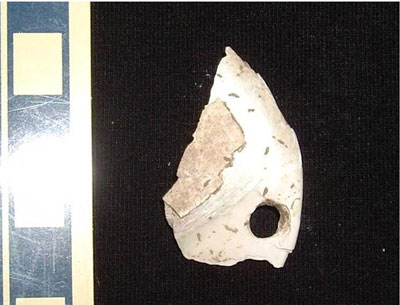
In all three features, there are many whole shells (Photo 1). There are also many shells that are almost whole, but are missing the ventral edges (Photo 2), and there are pieces that are only the ventral edges (Photo 3). Most of the nearly whole shells are relatively smooth along the broken edge, as shown in Photo 2, but some are uneven along the broken edge (Photo 4). There are only a few other fragments, some with hinges and some without. Shells in Photos 1-4 are from Feature D, but the same types of breakage are seen in Features C and SL.
Table 6. Whole shells and fragments of Gafrarium from Feature C.
Whole or fragment
NISP
Percent of NISP
MNI
Percent of MNI
Weight (grams)
Percent of Wt.
Whole shells
173
54.2
86.5
70.3
643.4
72.7
Whole shells with hewn perforations
2
0.6
1.0
0.8
5.9
0.7
Almost whole shells but ventral edges missing
59
18.5
29.5
24.0
149.3
16.9
Ventral edges
69
21.6
68.6
7.8
Other fragments, hinges
12
3.8
6.0
4.9
15.4
1.7
Other fragments without hinges
4
1.3
2.1
0.2
TOTALS
319
100.0
123
100.0
884.7
100.0
Table 7. Whole shells and fragments of Gafrarium from Feature D.
Whole or fragment
NISP
Percent of NISP
MNI
Percent of MNI
Weight (grams)
Percent of Wt.
Whole shells
114
9.2
57.0
21.2
416.7
25.6
Almost whole shells but ventral edges missing
387
31.1
193.5
71.9
688.3
42.4
Ventral edges
646
52.0
450.4
27.7
Other fragments, hinges present
37
3.0
18.5
6.9
28.8
1.8
Other fragments without hinges
59
4.7
39.6
2.4
TOTALS
1243
100.0
269
100.0
1623.8
100.0
Table 8. Whole shells and fragments of Gafrarium from Feature SL.
Whole or fragment
NISP
Percent of NISP
MNI
Percent of MNI
Weight (grams)
Percent of Wt.
Whole shells
31
40.3
15.5*
50.0
162.9
52.8
Almost whole shells, but ventral edges missing
32
41.6
16.0
50.0
126.2
40.9
Ventral edges
12
15.6
16.7
5.4
Other fragments without hinges
2
2.6
2.6
0.8
TOTALS
77
100.0
32.0
100.0
308.4
100.0
* counted as 16
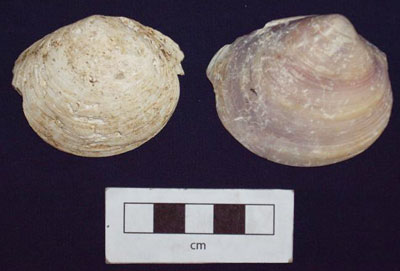
Photo 1. Whole Gafrarium shells from Feature D. Most are Gafrarium tumidum. On the basis of this photo, Dr. Katherine Szabó has suggested that the shell at the upper right is Gafrarium pectinatum.
Photo 2. Gafrarium shells from Feature D missing the ventral edges, but having relatively smooth broken edges.
Photo 3. Ventral edges of Gafrarium shells from Feature D.
Photo 4. Gafrarium shells from Feature D missing the ventral edges and having uneven breaks.
The three cultural features differ with regard to the percentage of whole shells of Gafrarium. Feature C has an MNI of 123 Gafrarium, which means there were 246 whole shells, and 70 percent of those 246 shells are still whole. Twenty-four percent of the 246 once-whole shells are almost whole but missing the ventral edges.
The opposite is true in Feature D. There are more shells missing the ventral edges than whole shells. The MNI of Feature D is 269, which means there were 538 whole shells. But only 21 percent of those 538 shells are now whole. Nearly 72 percent of the 538 shells are now missing the ventral edges.
Feature SL has an MNI of 32 Gafrarium, which means there were 64 whole shells. Now 48 percent of the shells are whole and 50 percent are missing the ventral edges. However the number of shells in Feature SL is small, so the percentages may not be very meaningful.
In addition to the shells missing ventral edges, there are two Gafrarium shells from Feature C with hewn perforations (Photo 5). The perforations appear to have been made by people, and they also appear to have been made rather recently. The perforations may have been made for the purpose of stringing the shells together or attaching the shells to something else.
Photo 5. Two Gafrarium shells from Feature C with hewn perforations.
The two grab samples do not exhibit the same breakage patterns as the cultural features. In the lagoon beach grab sample (Table 4), all of the Gafrarium shells are whole, and in the berm grab sample (Table 5), there are only two small fragments of Gafrarium.
Discussion
Habitat of Gafrarium tumidum
Baron and Clavier (1992) analyzed factors influencing the spatial distribution of Gafrarium tumidum on the southwest coast of New Caledonia, south and west of the Phoenix Islands. They found that depth was a more important factor than granulometric composition of the substrates. Maximum density and biomass of G. tumidum were found at the lower limit of areas uncovered by neap tides. It appears that the clams are limited by the neap low tides, that is, the highest low tides.
Gafrarium tumidum is found on a variety of substrates. Gibbs (1978) recorded G. tumidum from medium muddy sand of intertidal sand flats; Swadling and Chowning (1981) said this species is collected from muddy sediment pockets on the reef flat; Purchon and Purchon (1981) found G. tumidum in clean sand areas sheltered from waves and tidal currents. Baron and Clavier (1992) found maximum biomass in sediments comprising more than 50% medium, fine, and very fine sand, with 5 to 15% pebbles and granules, and at most 15% mud.
Baron and Clavier (1992) found that G. tumidum is comparatively independent of carbonate content, since high density and biomass were recorded with both very low (0 to 3%) and high (20 to 68%) carbonate contents.
The grab sample from the Nikumaroro lagoon beach shore yielded more Gafrarium shells than shells of any other genus, so it is evident that Gafrarium lives in the lagoon.
Both G. tumidum and G. pectinatum are considered common in the Indo-Pacific (Abbot and Dance 1983). Frank Thomas, who did his doctoral research on several atolls of the Gilbert Islands during the 1990s, has done excellent work on mollusk gathering in Western Kiribati linking behavioral ecology with ethnoarchaeology (Thomas 2002, 2007a, 2007b). Thomas’s articles do not mention G. tumidum, but do mention G. pectinatum, which is found in sand flat and seagrass habitats in Western Kiribati. Marshall Weisler, who has worked in the Marshall Islands, does not mention Gafrarium at Ujae Atoll (Weisler 1999).
Gafrarium was harvested both prehistorically and historically on Guam. Amesbury (2007) found that G. tumidum was more widespread in Guam during the early prehistoric period. It is now confined to inner Apra Harbor (Paulay 2003).
Food Value of Gafrarium
The food value of the Gafrarium clams in the three cultural features has been calculated in the following way. The weight of whole shells in a feature was divided by the number of whole shells to get the average weight per shell (Table 9). Then the average weight per shell was multiplied by two (because Gafrarium is a bivalve), and that result was multiplied by the MNI for the feature. In that way the estimated total shell weight of Gafrarium for each feature was determined.
Table 9. Estimated shell weight and meat weight in grams of all Gafrarium in Features C, D, and SL.
Feature
Weight of
whole shells (g)
Number of
whole shells
Average weight
per shell (g)
MNI
Estimated total
shell weight (g)
Estimated total
meat weight (g)
Estimated
number of
calories
C
643.4
173
3.72
123
915
431
315
D
416.7
114
3.66
269
1,969
927
677
SL
162.9
31
5.25
32
336
158
115
TOTALS
3,220
1,516
1,107
The weight of the meat per feature was calculated based on an experiment by Tom King discussed on the TIGHAR website:
“Earlier, I had procured a good-sized Cherrystone Clam from a local supermarket ($0.29 donation to TIGHAR). Its live, wet weight was found to be 136 grams. The clam was then boiled until it opened (about five minutes), and with some feeling of guilt the meat and innerds were removed and discarded. The wet empty shell weight of both valves was found to be 94 grams. The valves were then dried over a charcoal fire for two hours. The dry(er) weight was 92 grams.Note, this drying, on a rack about 15 cm. above the fire, did not cause cracking or any other noticeable change to the shell.”
In this particular Cherrystone clam, the dry shell accounted for 68% of the total weight, while the meat accounted for 32%.
A cherrystone clam (Mercenaria mercenaria) belongs to the family Veneridae, the same family to which Gafrarium belongs. Different common names are applied to different sizes of this species. According to Wikipedia, the common names in order of the smallest to the largest clams are countnecks, littlenecks, topnecks, cherrystones, and quahogs. According to Abbott and Dance (1983), the maximum length or diameter of an average adult specimen of M. mercenaria is 10 cm, while the same measurement for G. tumidum is 3.5 cm. The cherrystone in the experiment was a larger bivalve than a Gafrarium, but the assumption was made that the meat weight of Gafrarium is 32% of the total weight based on the above experiment.
The total estimated weight of Gafrarium meat in the three cultural features is 1,516 grams. According to Thomas (2007a), the number of calories per 100 grams of raw, wet edible weight of G. pectinatum is 73. Based on the information in Table 9 and assuming that G. tumidum has approximately the same food value as G. pectinatum, the Gafrarium in the cultural features provided approximately 1,107 calories.
Clams are a good source of protein and iron. The following website, Health Benefits of Clams, provides information on the nutritional value of 3 ounces (84.9 grams) of raw clams (no species named): Calories 63, Protein 10.9 g, Carbohydrate 2.2 g, Fat 0.8 g, Fiber 0.0 g, Iron 11.9 mg, and Vitamin B12 42 mcg.
Possible Explanations for the Breakage Patterns
The Introduction, Methods, and Results sections of this paper, including Photos 1-5, were sent electronically to Dr. Katherine Szabó, QEII Research Fellow at the University of Wollongong, Australia. Dr. Szabó is an expert on shell as a raw material for artifact production. She has provided a report (Appendix B: Notes on breakage patterns observed on Seven Site Gafrarium) with a diagram outlining possible causes of accidental breakage and deliberate breakage and visual examples of the various types of breakage.
Szabó states that “the valves with hewn perforations clearly represent deliberate human modification” but that the function of the modified valves is not clear (Szabó Appendix B). In a recent article about perforated Anadara shells from the Lapita site of Nukuleka on Tongatapu, Tonga, Connaughton et al. (2010) reviewed what other authors have said about the uses of perforated bivalves in the Pacific. Poulsen, who had worked at Nukuleka earlier, interpreted the perforated Anadara found there as net sinkers (Poulsen 1987). Spennemann (1993) reported that Anadara shells were ethnographically employed as net weights, vegetable scrapers, paring knives, coconut-grater heads, bark cloth scrapers, personal adornments, and parts of magic bundles. Spennemann acknowledged that removal of the umbo on an Anadara shell would make it possible to attach the shell to a net, but he also cautioned against over-interpretation, since accidental breakage and meat extraction might lead to a similar breakage pattern. Burly and Shutler (2007) disagreed with Spennemann based on the robusticity of Anadara and the lack of functional advantage in meat extraction of removing the umbo. They agreed with Poulsen that the perforated Anadara shells were net sinkers. Connaughton et al. (2010) said that some perforated Anadara shells may have served as net sinkers, but they proposed that many were by-products of a 3,000-year-old game.
Concerning the more common breakage pattern in which the ventral margins are broken from the bodies of the valves, Szabó suggests that the breakage may be natural or trampling breakage due in part to the small size of the Gafrarium shells, or it may be fatigue fracture from use of the shells as tools. The explanation for this type of breakage is as follows: “If a bivalve is used as a scraper or knife, then consistent downward pressure is being exerted on the ventral margin. With extended use, or the working of rough or difficult materials, the valve may snap during use” (Szabó Appendix B). Szabó suggests that confirmation of the use of the Gafrarium valves as tools would require close examination of the ventral margins. However, the use would have to have been heavy “for clear visual traces of use wear to be apparent” (Szabó Appendix B). High-power microscopy may be required.
Comparison with a Guam Shell Midden
The Nikumaroro features can be compared with a Gafrarium midden found about 2,000 feet inland of the west coast of Guam (Moore and Amesbury 2011). The midden was divided into six one-meter squares referred to as Units 1-6. Shells were collected from the surface of all units, and Unit 2 was excavated to 20 cm below surface where the midden ended. Weight of the shells from the surface collections of all six units is 1,804.6 grams, and weight of the shells from the excavation of Unit 2 is 11,518.7 grams. Gafrarium tumidum accounts for more than 80 percent of the weight of shells in the surface collections and more than 70 percent of the weight of shells from the excavation of Unit 2. A few historic items, including a fragment of an elephant figurine stamped “JAPAN,” a marble, a glass bead, some historic ceramic sherds, and some pieces of wire, nails and other rusty metal, found with the shells led the authors to estimate that the midden dates to the 1920s, 30s, or early 40s.
The breakage patterns in the Guam Gafrarium are similar to the Nikumaroro Gafrarium, except that there are no shells with hewn perforations. There are many whole shells and many shells with the ventral margins missing. Table 10 shows the average weight per shell of G. tumidum, and the percent of whole shells in the assemblage. The lightest weight shells were found at the surface, and the heaviest shells at the lowest depth. The surface assemblage has the lowest percentage of whole shells, and lowest level has the highest percentage of whole shells. The higher shells are more broken because they are lighter weight shells and/or because they are closer to the surface.
Table 10. Estimated weight per shell and percentage of whole shells of Gafrarium tumidum in all levels of Unit 2, Site 1, Piti, Guam (Moore and Amesbury 2011).
Level
Weight of whole shells (g)
Number of whole shells
Average weight per shell (g)
MNI
Percent of shells still whole
Surface
182.2
25
7.29
27
46%
0-10 cmbs
2653.8
298
8.91
266
56%
10-20 cmbs
3825.9
353
10.84
233
76%
TOTALS
6,661.9
676
9.85
526
64%
Table 11 shows the same kinds of information for the Nikumaroro features. The Nikumaroro shells are much lighter than the Guam shells. Average weight of the Nikumaroro shells is less than half the average weight of the Guam shells. Also there are fewer whole shells in the Nikumaroro features. Only 38 percent of all the Gafrarium are whole compared with 64 percent whole in the Guam assemblages (above). Feature D, which yielded the lightest shells, has only 21 percent of the shells whole. But Feature C, which yielded shells only slightly larger, has 70 percent of the shells whole, and Feature SL, which yielded the heaviest shells, has 48 percent whole.
Table 11. Estimated weight per shell and percentage of whole shells of Gafrarium tumidum in the three cultural features at Nikumaroro.
Feature
Weight of whole shells (g)
Number of whole shells
Average weight per shell (g)
MNI
Percent of shells still whole
Feature C
643.4
173
3.72
123
70%
Feature D
416.7
114
3.66
269
21%
Feature SL
162.9
31
5.25
32
48%
TOTALS
1,223.0
318
3.85
424
38%
It appears that Szabó is correct in thinking that the light weight of the Nikumaroro shells may explain why so many are broken. But the weight alone does not explain the data. It appears that something else is involved, since Feature C has 70 percent of the shells whole. It’s possible that Feature C was collected in a different manner from the other two features. It’s also possible that it is more recent, or that it was somehow protected from trampling. In an email dated 2/15/11, Tom King, archaeologist with TIGHAR, stated, “The map also doesn’t show how asphalt siding had been rolled out on the ground, but it clearly had; we observed its residual flecks all through the area overlying Gafrarium Feature C.” Maybe the asphalt siding protected Feature C from breakage.
Recommendations
It was anticipated that the study of the shells would help to answer the question of who collected the shells, as well as how the shells were used. Some of the possible groups of people are 1) pre-twentieth century Pacific Islanders, 2) Euro-American castaways, such as Amelia Earhart and Fred Noonan, and 3) twentieth century Pacific Islanders, such as the settlers associated with the Phoenix Island Settlement Scheme (see King et al. 2001). The study so far has not ruled out any of these three groups.
In addition to examining the ventral margin fragments of the shells with a high-powered microscope for traces of use-wear, the author and Dr. Szabó recommend experimental work with Gafrarium in order to discover how the ventral margins were separated from the valves. Using Gafrarium valves as tools for peeling taro, scraping coconut, cutting bamboo, and other likely activities may duplicate the breakage pattern seen in the shells from the cultural features.
Other recommended work would include trampling/compaction loading experiments following methods outlined in paleontology. If these experiments produce the separation of the ventral margins from the valves, then we might wonder if the logging operation which took place on Nikumaroro about 1940 (see Gillespie 2001:5) was responsible for the breakage pattern.
Live Gafrarium should also be collected and both raw and cooked specimens opened with a jackknife. If prying open the shells in that way separates the ventral margins, that would point to a Euro-American castaway. It is possible that the three cultural features represent three meals for one or two people. The differences in frequencies of the broken valves may be due to a castaway becoming more proficient at opening the shells without breaking them or, on the other hand, becoming more desperate to obtain food and not caring if the shells break. It is doubtful that we could determine the order in which the three cultural features were formed.
Shells from the cultural features could be radiocarbon dated. However, the date of the disappearance of Amelia Earhart and Fred Noonan (1937) and the date of arrival of the first work party associated with the Phoenix Island Settlement Scheme (1938) are too close for radiocarbon dating to discriminate between the two events. Dating the shells might reveal if the cultural features were created by pre-twentieth century Pacific Islanders.
References
Abbott, R. Tucker and S. Peter Dance. 1983.
Compendium of Seashells. E.P. Dutton, Inc., New York.
Amesbury, Judith R. 2007.
Mollusk collecting and environmental change during the Prehistoric Period in the Mariana Islands. Coral Reefs 26:947-958.
Baron, Jérôme and Jacques Clavier. 1992.
Effects of environmental factors on the distribution of the edible bivalves Atactodea striata, Gafrarium tumidum and Anadara scapha on the coast of New Caledonia (SW Pacific). Aquat. Living Resour., 5(2), 107-114.
Burley, D.V. and R. Shutler, Jr. 2007.
Ancestral Polynesian fishing gear: Archaeological insights from Tonga. In A. Anderson, K. Green, and F. Leach (eds), Vastly Ingenious: The Archaeology of Pacific Material Culture in Honour of Janet M. Davidson, pp. 155-172. Otago University Press, Otago.
Connaughton, Sean P., Karine Taché, and David V. Burley. 2010.
Taupita: A 3000-year-old shell game in the Lapita cultural complex of Tonga. Journal of Social Archaeology 10(1): 118-137.
Gibbs, P.E. 1978.
Macrofauna of the intertidal sand flats on low wooded islands, northern Great Barrier Reef. Phil. Trans. R. Soc. Lond. 283, 81-97.
Gillespie, Ric. 2001.
TIGHAR in Tarawa. TIGHAR Tracks 17(3): 1-7. TIGHAR, Wilmington Delaware.
Archaeological Investigation of Site 66-03-1659, Lot 170-1NEW-R5, Piti, Guam. Prepared for Joseph D. Lopez and Robert P. Salas, Landscape Management Systems, Inc. Micronesian Archaeological Research Services, Inc., Mangilao, Guam.
Paulay, Gustav. 2003.
Marine Bivalvia (Mollusca) of Guam. Micronesica 35-36:218-243.
Poulsen, J. 1987.
Early Tongan Prehistory: The Lapita Period on Tongatapu and its Relationships. Terra Australis 12. Pandanus Books, Australian National University, Canberra.
Purchon R.D. and D.E.A. Purchon. 1981.
The marine shelled Mollusca of West Malaysia and Singapore. I. General introduction and an account of the collections. J. Moll. Stud., 47, 290-312.
Spenneman, D.H.R. 1993.
Ark shell netsinkers: Fact or fiction. Archaeology in New Zealand 36:75-83.
Swadling, P. and A. Chowning. 1981.
Shellfish gathering at Nukalau island, West New Britain province, Papua New Guinea. J. Soc. Océanistes, 38, 159-167.
Thomas, F.R. 2002.
An evaluation of central-place foraging among mollusk gatherers in Western Kiribati, Micronesia: Linking behavioral ecology with ethnoarchaeology. World Archaeology Vol. 34(1), 182-208.
Thomas, F.R. 2007a.
The behavioral ecology of shellfish gathering in Western Kiribati, Micronesia. 1: Prey choice. Human Ecology 35, 179–194.
Thomas, F.R. 2007b.
The behavioral ecology of shellfish gathering in Western Kiribati, Micronesia. 2: Patch choice, patch sampling, and risk. Human Ecology 35: 515-526.
Weisler, M.I. 1999.
Atolls as settlement landscapes: Ujae, Marshall Islands. Atoll Research
Bulletin No. 460.
National Museum of Natural History, Smithsonian Institution, Washington, D.C.
APPENDIX A
Counts and Weights of Shells from the Seven Site, Nikumaroro
By Judith R. Amesbury
Micronesian Archaeological Research Services, Guam
Key:
Wh. #
=
Number of whole shells
Fr. #
=
Number of shell fragments
Wt. (g)
=
Weight in grams
sp.
=
unidentified species
spp.
=
more than one unidentified species
Table 1. Number and weight in grams of whole shells and fragments from Feature C, 2001 collection.
FAMILY
GENUS
SPECIES
Wh. #
Wt. (g)
Fr. #
Wt. (g)
Total Wt.
Percent by Wt.
Veneridae
Gafrarium
tumidum
81
284.2
22
55.7
339.9
99.65
Bivalves
81
284.2
22
55.7
339.9
99.65
Cypraeidae
Cypraea
sp.
1
1.2
1.2
0.35
Gastropods
1
1.2
1.2
0.35
TOTALS
81
284.2
23
56.9
341.1
100.00
Table 2. Number and weight in grams of whole shells and fragments from Feature C, 2010 collection.
FAMILY
GENUS
SPECIES
Wh. #
Wt. (g)
Fr. #
Wt. (g)
Total Wt.
Percent by Wt.
Tellinidae
Quidnipagus
palatam
1
1.0
1.0
0.16
Tridacnidae
Tridacna
maxima
1
45.2
45.2
7.44
Veneridae
Gafrarium
tumidum
92
359.2
124
185.6
544.8
89.68
Bivalves
93
404.4
125
186.6
591.0
97.28
Conidae
Conus
sp.
1
2.6
2.6
0.43
Turbinidae
Turbo
sp.
4
13.9
13.9
2.29
Gastropods
5
16.5
16.5
2.72
TOTALS
93
404.4
130
203.1
607.5
100.00
Table 3. Number and weight in grams of whole shells and fragments from Feature D, Bag 1 of 2, collected 6/2/10.
FAMILY
GENUS
SPECIES
Wh. #
Wt. (g)
Fr. #
Wt. (g)
Total Wt.
Percent by Wt.
Cardiidae
Fragum
fragum
37
11.1
11.1
1.35
Mytilidae
1
0.1
0.1
0.01
Veneridae
Gafrarium
tumidum
30
104.8
641
555.6
660.4
80.10
Bivalves
67
115.9
642
555.7
671.6
81.46
Cerithiidae
9.7
1.18
Cerithium
nodulosum
1
5.8
Other
spp. – at least 2 other species
2
0.5
5
3.4
Conidae
Conus
spp. – at least 3 species
6
20.9
4
4.5
25.4
3.08
Cypraeidae
Cypraea
spp. – at least 3 species
12
54.7
15
51.5
106.2
12.88
Muricidae
2
3.4
3.4
0.41
Neritidae
Nerita
sp.
1
0.5
0.5
0.06
Strombidae
2
1.6
1.6
0.19
Turbinidae
Turbo
sp.
4
6.1
6.1
0.74
Gastropods
20
76.1
34
76.8
152.9
18.54
TOTALS
87
192.0
676
632.5
824.5
100.0
Table 4. Number and weight in grams of whole shells and fragments from Feature D, Bag 2 of 2, collected 6/3/10.
FAMILY
GENUS
SPECIES
Wh. #
Wt. (g)
Fr. #
Wt. (g)
Total Wt.
Percent by Wt.
Cardiidae
Fragum
fragum
8
2.6
2.6
0.25
Veneridae
Gafrarium
tumidum
84
311.9
488
651.5
963.4
91.44
Bivalves
92
314.5
488
651.5
966.0
91.69
Conidae
Conus
sp.
2
25.0
25.0
2.37
Cypraeidae
Cypraea
spp. – at least 2 species
8
48.2
3
8.4
56.6
5.37
Turbinidae
Turbo
sp.
1
6.0
6.0
0.57
Gastropods
10
73.2
4
14.4
87.6
8.31
TOTALS
102
387.7
492
665.9
1053.6
100.00
Table 5. Number and weight in grams of whole shells and fragments from Feature SL.
FAMILY
GENUS
SPECIES
Wh. #
Wt. (g)
Fr. #
Wt. (g)
Total Wt.
Percent by Wt.
Cardiidae
Fragum
fragum
2
0.8
0.8
0.24
Tridacnidae
Tridacna
maxima
1
5.6
5.6
1.68
Veneridae
321.0
96.02
Gafrarium
tumidum
31
162.9
46
145.5
Pitar
prora
2
12.6
Bivalves
36
181.9
46
145.5
327.4
97.94
Turbinidae
Turbo
sp.
2
6.8
6.8
2.03
Unidentified gastropods
1
0.1
0.1
0.03
Gastropods
3
6.9
6.9
2.06
TOTALS
36
181.9
47
152.4
334.3
100.00
Table 6. Number and weight in grams of whole shells and fragments from the lagoon beach grab sample.
FAMILY
GENUS
SPECIES
Wh. #
Wt. (g)
Fr. #
Wt. (g)
Total Wt.
Percent by Wt.
Arcidae
Barbatia
cf.amygdalumtostum
1
5.2
5.2
1.06
Cardiidae
Fragum
fragum
1
0.2
1
2.2
2.4
0.49
Chamidae
Chama
cf.iostoma
3
36.0
36.0
7.37
Lucinidae
Epicodakia
bella
2
4.1
4.1
0.84
Tellinidae
Quidnipagus
palatam
1
3.8
3.8
0.78
Tridacnidae
222.6
45.57
Tridacna
maxima
4
46.4
2
51.9
Tridacna
spp. (at least 2)
3
124.3
Veneridae
214.4
43.89
Gafrarium
tumidum
28
120.6
Pitar
prora
10
78.1
1
2.0
Lioconcha
cf.castrensis
2
13.7
Bivalves
52
308.1
7
180.4
488.5
100.00
Gastropods
0
0
0
0
0
0.0
TOTALS
52
308.1
7
180.4
488.5
100.00
Table 7. Number and weight in grams of whole shells and fragments from the grab sample from the berm that fronts the lagoon shore.
FAMILY
GENUS
SPECIES
Wh. #
Wt. (g)
Fr. #
Wt. (g)
Total Wt.
Percent by Wt.
Cardiidae
Fragum
fragum
333
63.0
19
2.8
65.8
82.46
Tellinidae
probably 2 species
2
0.2
0.2
0.25
Veneridae
Gafrarium
tumidum
2
0.5
0.5
0.63
Unidentified bivalves
10
1.0
1.0
1.25
Bivalves
333
63.0
33
4.5
67.5
84.59
Cerithiidae
at least 2 species
41
7.8
27
3.8
11.6
14.53
Strombidae
Strombus
sp.
1
0.7
0.7
0.88
Gastropods
41
7.8
28
4.5
12.3
15.41
TOTALS*
374
70.8
61
9.0
79.8
100.00
* Included with the shells on this table are 169 small pieces of coral weighing 23.6 grams. In addition to the above shells and the accompanying 23.6 grams of coral, there are 50 grams of unsorted shells and coral pieces with approximately the same make-up as the shells and corals that were counted and weighed. The total weight of shells and coral fragments from the berm grab sample is therefore 153.4 grams.
APPENDIX B
Notes on Breakage Patterns Observed on Seven Site Gafrarium
By Dr. Katherine Szabó, QEII Research Fellow
School of Earth and Environmental Sciences
University of Wollongong, Australia
Shell breakage can be caused through a number of processes, with breakage patterns being strongly influenced by the macro- and microstructure of the species being considered (Szabó 2008). Whilst far from exhaustive, the following outlines potential causes of breakage in Gafrarium spp. valves at the Seven Site, and what the traces of these processes are likely to be.
Gafrarium spp. macro- and microstructure
All bivalves within the Veneridae have a dominantly crossed lamellar microstructure as is typical of the most evolved bivalve families. This means that bundles of aragonite crystals are aligned at 45° angles, with the effect that any force delivered to the surface of the shell will tend to travel a zigzag path following the line of, but not breaking, the crystal stacks. Thus, breakage in crossed lamellar structures has a much more „ragged‟ appearance than, for example, the shearing of lamellar sheets seen in nacreous (mother-of-pearl) structures.
Compared to many other bivalve families, venerids are also relatively robust and incidences of in situ breakage are less common than that seen in families such as the Mytilidae (mussels), Ostreidae (oysters) or Tellinidae (tellins). Thus, the high incidence of Gafrarium spp. breakage at the Seven Site is worthy of note and warrants explanation.
Causes of breakage
Visual Examples
Thermal fracture
Tridacna gigas (modern midden, Pulau Seraya, Flores, Indonesia, August 2010)
Shells with deliberate hewn perforations
Trochus niloticus fragment from the earliest Lapita site in Fiji, Bourewa
Polymesoda erosa valves with hewn perforations from the Kain Hitam burial site, Niah Caves, Sarawak, Malaysia. Reference: Szabó et al. 2008
One of a number of Codakia punctata valves with central, hewn perforations. Bourewa Lapita site, Fiji.
One of a number of Fimbria fimbriata valves with hewn perforations. Bourewa Lapita site, Fiji.
Fatigue fracture from tool use
Batissa violacea valves with the ventral margin snapped away. This was typical of B. violacea breakage at this site. Holocene (H. sapiens) levels of Liang Bua, Flores, Indonesia.
Snap fracture surface on a Batissa violacea valve at 10x magnification. The fracture is rather smooth at the macro-level, but rough at the micro-level. Liang Bua, Flores, Indonesia.
Ventral margin with use-wear, Batissa violacea. Liang Bua Holocene levels, Flores, Indonesia.
A note on interpretation following experimental work
In 2010, Szabó supervised BA(hons) student Brent Koppel in a series of experiments investigating diagnostic traces of use-wear in limpets (Koppel 2010). The results of this study are currently being prepared for publication. In this study, limpets in the families Nacellidae, Patellidae and the “false limpet” family Siphonariidae were used to peel taro corms and yam tubers, scrape bamboo, cut pork flesh, scrape fresh pig bone and scrape haematite nodules to produce powdered pigment. The results were somewhat unexpected. Limpets transpired to be very effective peeling tools, but even after 100 scrapes of the shell against the taro/yam, few if any visible traces of wear were present on the margin. The most surprising result came from the haematite scraping experiment. Not only was a mound of powdered pigment produced in a short time, but even under a scanning electron microscope at 400x magnification the only traces of working were traces of red pigment on the margins of the shells.
While a number of important conclusions were drawn from this study, the two most significant are that, (1) shell can be a much more effective raw material for tool use than commonly assumed, and (2) visible use-wear traces are the product of extensive use, with casual or short term usage often leaving no visible traces at all.
Observations on breakage patterns of the Seven Site Gafrarium
Two types of breakage are manifest in the Seven Site Gafrarium samples: (1) valves with hewn perforations, and (2) valves where the ventral margin has come away from the body of the valve. In my experience, the first type is not natural breakage for Gafrarium spp., but the second type may be natural or trampling breakage.
Hewn perforations
From the photographs, the valves with hewn perforations clearly represent deliberate human modification. Whether “net-sinkers,” debris from game-playing, or any one of a number of functions is not clear and will likely remain so without detailed experimentation and investigation of use-wear.
Broken valves
Given the general robusticity of Gafrarium valves, the consistent levels of breakage across the samples from the Seven Site are unusual. However it should be noted that the average size of the valves is very small, and thus the valves are more fragile than is usual for Gafrarium in archaeological samples. The noted breakage may be natural or trampling breakage, or it may be fatigue fracture from using the shells as tools. I have never seen this particular pattern of breakage at such levels on Gafrarium valves, but have seen it on the robust valves of brackish-water clams in the Corbiculidae (Polymesoda spp., Batissa violacea). These species are widely reported as tools across the Asia-Pacific region, performing cutting and scraping tasks. Confirmation of the use of unmodified Gafrarium valves as tools would require close examination of the ventral margin fragments. However, given the results of the limpet study cited above, use would have to be heavy for clear visual traces of use-wear to be apparent.
References
Koppel, B. 2010.
Fracture and modification in Patella shells: isolating human working. Unpublished BA(hons) thesis. Centre for Archaeological Science, University of Wollongong.
Szabó, K. 2008.
Shell as a Raw Material: Mechanical Properties and Working Techniques. Archaeofauna 17: 125-138.
Szabó, K., Piper, P., Barker, G. 2008.
Sailing between worlds: the symbolism of death in northwest Borneo. In G. Clark, S. O’Connor and B.F. Leach (eds.) Islands of Inquiry, pp. 149-170. Canberra, ANU E-Press.
Copyright 2021 by TIGHAR, a non-profit foundation.
No portion of the TIGHAR Website may be reproduced by xerographic,
photographic, digital or any other means for any purpose. No portion
of the TIGHAR Website may be stored in a retrieval system, copied,
transmitted or transferred in any form or by any means, whether electronic,
mechanical, digital, photographic, magnetic or otherwise, for any purpose
without the express, written permission of TIGHAR. All rights reserved.





 Photo 1. Whole Gafrarium shells from Feature D. Most are Gafrarium tumidum. On the basis of this photo, Dr. Katherine Szabó has suggested that the shell at the upper right is Gafrarium pectinatum.
Photo 1. Whole Gafrarium shells from Feature D. Most are Gafrarium tumidum. On the basis of this photo, Dr. Katherine Szabó has suggested that the shell at the upper right is Gafrarium pectinatum.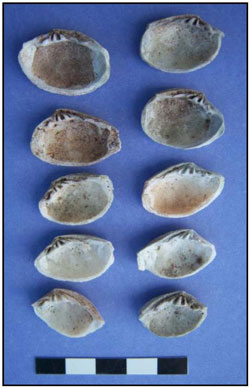 Photo 2. Gafrarium shells from Feature D missing the ventral edges, but having relatively smooth broken edges.
Photo 2. Gafrarium shells from Feature D missing the ventral edges, but having relatively smooth broken edges.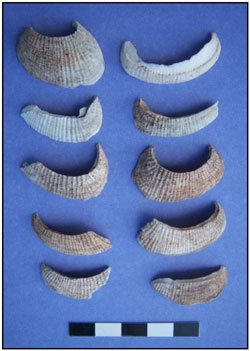 Photo 3. Ventral edges of Gafrarium shells from Feature D.
Photo 3. Ventral edges of Gafrarium shells from Feature D. Photo 4. Gafrarium shells from Feature D missing the ventral edges and having uneven breaks.
Photo 4. Gafrarium shells from Feature D missing the ventral edges and having uneven breaks.